Zasadniczo uważam rybie mózgi za raczej mało interesujące. Zbyt prosty to krewny mózgów ssaczych, zbyt „niedinozaurzy” – mózgów dinozaurów. Mózgi rekinów i okoni załapują się na odrobinę uwagi podczas zajęć z anatomii porównawczej, ale też głównie przy akompaniamencie lamentów nad tym jak bardzo są one prymitywne. Odhaczmy je na liście, zróbmy szkice, zaliczmy sprawdzian i przejdźmy dalej, by dowiedzieć się, jak rybie mózgi wyewoluowały w coś ciekawszego.
Ale zupełnie niedawno dowiedziałem się, że bezwzględność mojego podejścia jest bardziej nawet niż nieco niesprawiedliwa. Otóż Alice Clement i Per Ahlberg z Uniwersytetu w Uppsali (Szwecja) opublikowali w zeszłym tygodniu (koniec listopada 2014, przyp. tłum.) naprawdę fascynującą pracę – rzut oka na rekonstrukcję mózgu pochodzącej sprzed 380 milionów lat australijskiej ryby dwudysznej (Rhinodipterus). W zestawieniu z wcześniejszymi publikacjami wyłania się z niej niezwykle skomplikowany obraz ewolucji mózgu dwudysznych.
Dwudyszne są szczególnie interesującym obiektem badań nie tylko ze względu na ich relatywnie długą historię zapisu kopalnego (ponad 400 milionów lat), ale też z powodu ich kluczowej dla ewolucji kręgowców pozycji. Obecnie wyróżniamy trzy rodzaje ryb dwudysznych – Neoceratodus (rogoząb australijski), Lepidosiren (prapłetwiec południowoamerykański) i Protopterus (zamieszkujące Afrykę prapłetwce). Jedynie ten ostatni liczy sobie więcej niż jeden gatunek. Dwudyszne są prawdopodobnie rybami najbliżej spokrewnionymi z wyposażonymi w kończyny kręgowcami (tetrapodami, czyli czworonogami, w tym salamandrami, indykami i ludmi, by wymienić choć kilka przykładów), są zatem istotnym punktem odniesienia dla zrozumienia ich ewolucji.
Problem z badaniami mózgu dwudysznych to kwestia dwupoziomowa. Po pierwsze mózgi zazwyczaj w niewielkim stopniu podlegają fosylizacji, paleontolodzy muszą więc nieco oszukiwać, by móc przyjrzeć się mózgom wymarłych zwierząt. Mózgowie zamknięte jest w obrębie puszki mózgowej, struktury czaszki, która często zachowuje przynajmniej zgrubne jego kontury. Skorzystaj z tomografu komputerowego (TK), by obejrzeć wnętrze puszki mózgowej, a będziesz w stanie wykonać cyfrowy model mózgu, ale…. Cóż, puszka mózgowa – zarówno współczesnych dwudysznych, jak i licznych ich kopalnych krewniaków – jest w znacznej mierze strukturą chrzęstną, zatem nawet podstawy anatomii mózgu są nieomalże niepoznawalne w przypadku sporej części linii ewolucyjnej dwudysznych.
Na szczęście wiele wczesnych dwudysznych miało jednak skostniałe czaszki. Nadal jednak liczne spośród znanych szczątków to obiekty zmiażdżone, zdekompletowane bądź też jedynie częściowo zbadane. Szczęśliwie wszakże dysponujemy skamieniałościami z australijskiego rejonu Gogo, formacji skalnej sprzed 380 milionów lat obejmującej pozostałości rafy koralowej. Pochodzące stamtąd preparaty bywają naprawdę spektakularne (chociażby słynnyrybi zarodek połączony z rybią „matką” pępowiną), dobrze zachowane, ładnie oddzielające się od ścian swych skalnych kołysek przy użyciu odpowiednich metod. Jedna z tamtejszych skamieniałości to właśnie częściowy szkielet dwudysznej ryby z gatunku Rhinodipterus kimberleyensis. Jej puszka mózgowa została przepuszczona przez TK, co pozwoliło na opracowanie cyfrowego modelu wewnętrznych konturów czaszki i przybliżonych – mózgu. Ten z kolei model porównano z opublikowanymi dotąd danymi na temat mózgowia obecnie żyjących dwudysznych i dwudysznych kopalnych.
Jedną z najbardziej uderzających obserwacji jest ta, iż zrekonstruowany mózg Rhinodipterus kimberleyensis znacznie bardziej przypomina budową narząd rogozęba australijskiego (ostatnia ilustracja artykułu) niż pozostałych współczesnych dwudysznych (tych z Afryki i Ameryki Południowej, z rodzajów Lepidosiren i Protopterus). Jako że ich mózgowia mają podobny, dość prymitywny kształt, Clement i Ahlberg wysnuli hipotezę, że ukształtowanie mózgu pozostałych współczesnych dwudysznych wyewoluowało jako zupełnie unikalna, odrębna innowacja ewolucyjna. Inni badacze odnotowali już wcześniej, że mózgi ryb rodzajów Lepidosiren i Protopterus wykazują pewne podobieństwa do mózgów dzisiejszych płazów, jeśli jednak przyjrzeć się całości drzewa ewolucyjnego, jest niemalże oczywiste, że to tylko efekt konwergencji.
Clement i Ahlberg poświęcili też chwilę rozważaniom jak budowa mózgu przełożyła się na jego funkcje i zachowania zwierząt. Zauważyli, że w toku swej ewolucji dwudyszne najprawdopodobniej wzmocniły znacząco zmysł węchu, co można wnioskować ze zmian w wielkości i kształcie części mózgowia zań odpowiedzialnej. Pewne różnice w budowie ucha wewnętrznego (związanego nie tylko z odbiorem dźwięku, ale też z rejestrowaniem zmian ułożenia ciała i ruchu) są także przy porównaniu gatunków dość ewidentne, choć tu już konkretne implikacje zmian są nieco bardziej mgliste.
Mózgi dwudysznych są jednak zaskakująco interesujące! Muszę zresztą pogratulować autorom pracy wyjątkowo przystępnego tekstu. Skoro paleontolog od dinozaurów jest w stanie go zrozumieć, to to już jest pewne osiągnięcie! Wciąż, oczywiście, nawet z tym nowym badaniem, pozostaje jeszcze wiele do nauczenia się. Pozostaje mieć nadzieję, że badania innych dobrze zachowanych preparatów dostarczą nam dalszych informacji o tym, co siedzi w łebkach tych fascynujących zwierzaków.
Literatura:
Clement AM, Ahlberg PE (2014) The first virtual cranial endocast of a lungfish (Sarcopterygii: Dipnoi). PLOS ONE 9(11): e113898. doi:10.1371/journal.pone.0113898
2. Kopalna przedstawicielka dwudysznych z rodzaju Dipterus, bliska krewna Rhinodipterus kimberleyensis z pracy Clement & Ahlberga (2014); ilustracja z Ray 1907, domena publiczna.
3. Odlew cyfrowy i jego rysunek interpretacyjny – Rhinodipterus. Przód mózgu znajduje się z prawej strony, tył (rdzeń kręgowy) z lewej. Obszar ucha wewnętrznego wybarwiono na pomarańczowo, tyłomózgowie (wraz z podstawą rdzenia kręgowego) na żółto, śródmózgowie na niebiesko, przodomózgowie na czerwono i zielono; zmodyfikowana wersja Clement & Ahlberg 2014, CC-BY.
Ból- skandal czy błogosławieństwo? Postawy wobec bólu a ogólna wizja świata i człowieka (na przykładzie ateistów, buddystów i katolików) Sylwia Wojas Instytut…
Dostrzegam w naszym kraju pewną tendencję do zamierania dyskusji i polemiki między reprezentantami równych ideologii politycznych. Jako liberał i eks-konserwatysta jestem tym faktem dość zaniepokojony,…
Ponieważ w naszym kraju trwa zalew prawicowych ksiąg „mądrości” politycznej, i nikt z normalnych liberałów (a nie konserwatywnych wolnorynkowców, którzy mienią się być liberałami), nie…
Indie to kraj i cywilizacja zarazem, postrzegane są przez Zachód jako kraina baśni i mitów; w końcu hinduizm zakłada istnienie dziesiatków milionów bogów i demonów. …

 Problem z badaniami mózgu dwudysznych to kwestia dwupoziomowa. Po pierwsze mózgi zazwyczaj w niewielkim stopniu podlegają fosylizacji, paleontolodzy muszą więc nieco oszukiwać, by móc przyjrzeć się mózgom wymarłych zwierząt. Mózgowie zamknięte jest w obrębie puszki mózgowej, struktury czaszki, która często zachowuje przynajmniej zgrubne jego kontury. Skorzystaj z tomografu komputerowego (TK), by obejrzeć wnętrze puszki mózgowej, a będziesz w stanie wykonać cyfrowy model mózgu, ale…. Cóż, puszka mózgowa – zarówno współczesnych dwudysznych, jak i licznych ich kopalnych krewniaków – jest w znacznej mierze strukturą chrzęstną, zatem nawet podstawy anatomii mózgu są nieomalże niepoznawalne w przypadku sporej części linii ewolucyjnej dwudysznych.
Problem z badaniami mózgu dwudysznych to kwestia dwupoziomowa. Po pierwsze mózgi zazwyczaj w niewielkim stopniu podlegają fosylizacji, paleontolodzy muszą więc nieco oszukiwać, by móc przyjrzeć się mózgom wymarłych zwierząt. Mózgowie zamknięte jest w obrębie puszki mózgowej, struktury czaszki, która często zachowuje przynajmniej zgrubne jego kontury. Skorzystaj z tomografu komputerowego (TK), by obejrzeć wnętrze puszki mózgowej, a będziesz w stanie wykonać cyfrowy model mózgu, ale…. Cóż, puszka mózgowa – zarówno współczesnych dwudysznych, jak i licznych ich kopalnych krewniaków – jest w znacznej mierze strukturą chrzęstną, zatem nawet podstawy anatomii mózgu są nieomalże niepoznawalne w przypadku sporej części linii ewolucyjnej dwudysznych. Jedną z najbardziej uderzających obserwacji jest ta, iż zrekonstruowany mózg Rhinodipterus kimberleyensis znacznie bardziej przypomina budową narząd rogozęba australijskiego (ostatnia ilustracja artykułu) niż pozostałych współczesnych dwudysznych (tych z Afryki i Ameryki Południowej, z rodzajów Lepidosiren i Protopterus). Jako że ich mózgowia mają podobny, dość prymitywny kształt, Clement i Ahlberg wysnuli hipotezę, że ukształtowanie mózgu pozostałych współczesnych dwudysznych wyewoluowało jako zupełnie unikalna, odrębna innowacja ewolucyjna. Inni badacze odnotowali już wcześniej, że mózgi ryb rodzajów Lepidosiren i Protopterus wykazują pewne podobieństwa do mózgów dzisiejszych płazów, jeśli jednak przyjrzeć się całości drzewa ewolucyjnego, jest niemalże oczywiste, że to tylko efekt konwergencji.
Jedną z najbardziej uderzających obserwacji jest ta, iż zrekonstruowany mózg Rhinodipterus kimberleyensis znacznie bardziej przypomina budową narząd rogozęba australijskiego (ostatnia ilustracja artykułu) niż pozostałych współczesnych dwudysznych (tych z Afryki i Ameryki Południowej, z rodzajów Lepidosiren i Protopterus). Jako że ich mózgowia mają podobny, dość prymitywny kształt, Clement i Ahlberg wysnuli hipotezę, że ukształtowanie mózgu pozostałych współczesnych dwudysznych wyewoluowało jako zupełnie unikalna, odrębna innowacja ewolucyjna. Inni badacze odnotowali już wcześniej, że mózgi ryb rodzajów Lepidosiren i Protopterus wykazują pewne podobieństwa do mózgów dzisiejszych płazów, jeśli jednak przyjrzeć się całości drzewa ewolucyjnego, jest niemalże oczywiste, że to tylko efekt konwergencji.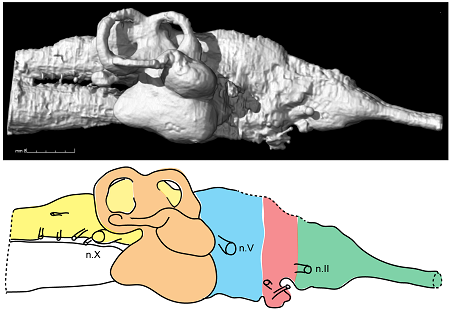 Mózgi dwudysznych są jednak zaskakująco interesujące! Muszę zresztą pogratulować autorom pracy wyjątkowo przystępnego tekstu. Skoro paleontolog od dinozaurów jest w stanie go zrozumieć, to to już jest pewne osiągnięcie! Wciąż, oczywiście, nawet z tym nowym badaniem, pozostaje jeszcze wiele do nauczenia się. Pozostaje mieć nadzieję, że badania innych dobrze zachowanych preparatów dostarczą nam dalszych informacji o tym, co siedzi w łebkach tych fascynujących zwierzaków.
Mózgi dwudysznych są jednak zaskakująco interesujące! Muszę zresztą pogratulować autorom pracy wyjątkowo przystępnego tekstu. Skoro paleontolog od dinozaurów jest w stanie go zrozumieć, to to już jest pewne osiągnięcie! Wciąż, oczywiście, nawet z tym nowym badaniem, pozostaje jeszcze wiele do nauczenia się. Pozostaje mieć nadzieję, że badania innych dobrze zachowanych preparatów dostarczą nam dalszych informacji o tym, co siedzi w łebkach tych fascynujących zwierzaków.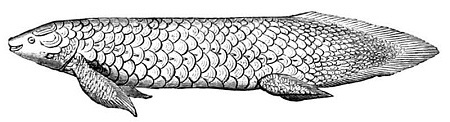
{kind=link}
{kind=link}



